


In Chapter 1, we explored the importance of building synthetic organelles, particularly in bacteria. From early efforts with fusion proteins and molecular scaffolds to creating more complex cellular structures, the idea of spatial organization in bacterial cells is a game-changer for biotechnology. However, attempts to construct membrane-bound organelles have been limited by our incomplete understanding of their biogenesis.
Fortunately, nature provides an exciting alternative to membrane-bound compartments - membraneless organelles. Let’s dive into the story of membraneless organelles, from their early discovery to modern efforts in understanding their formation, which guide us toward building synthetic versions.
What Are Membraneless Organelles? A Peek into History
Using the building metaphor, membrane-bound organelles are like rooms with walls, clearly separated from the rest of the space. In contrast, membraneless organelles are more like an open kitchen, condensed with food and materials. While they have no clearly defined barrier, they still block irrelevant elements from the outside and allow foods to move in and out as needed.
Many might think membraneless organelles are a "new discovery", especially given the attention they've received in recent years. In reality, the discovery of membraneless organelles goes back to the days of early microscopy.
Since Robert Hooke first observed and coined the term "cells," scientists in the 19th century began using optical microscopes to study cells. This work laid the foundation for the "cell theory", which became central to modern biology. Subcellular structures were identified and described early on around the same time:
- Nucleus, the first described organelle, by Antonie van Leeuwenhoek (1632–1723), Franz Bauer (1804) and Robert Brown (1831)1.
- Nucleolus, documented by Wagner (1835) and Valentin (1836-1839)2.
- Chloroplasts, observed by Antonie van Leeuwenhoek (1678) and later described as “chlorophyll granules” by Hugo von Mohl (1837)3.
- Mitochondrion, identified by Albert von Kolliker in 18574.
- Golgi apparatus, discovered by Camillo Golgi in 18985.
- Ergastoplasm, identified by Charles Garnier in 18976, later renamed as Endoplasmic Reticulum (ER), by Keith R Porter in 1953 (see below)7.
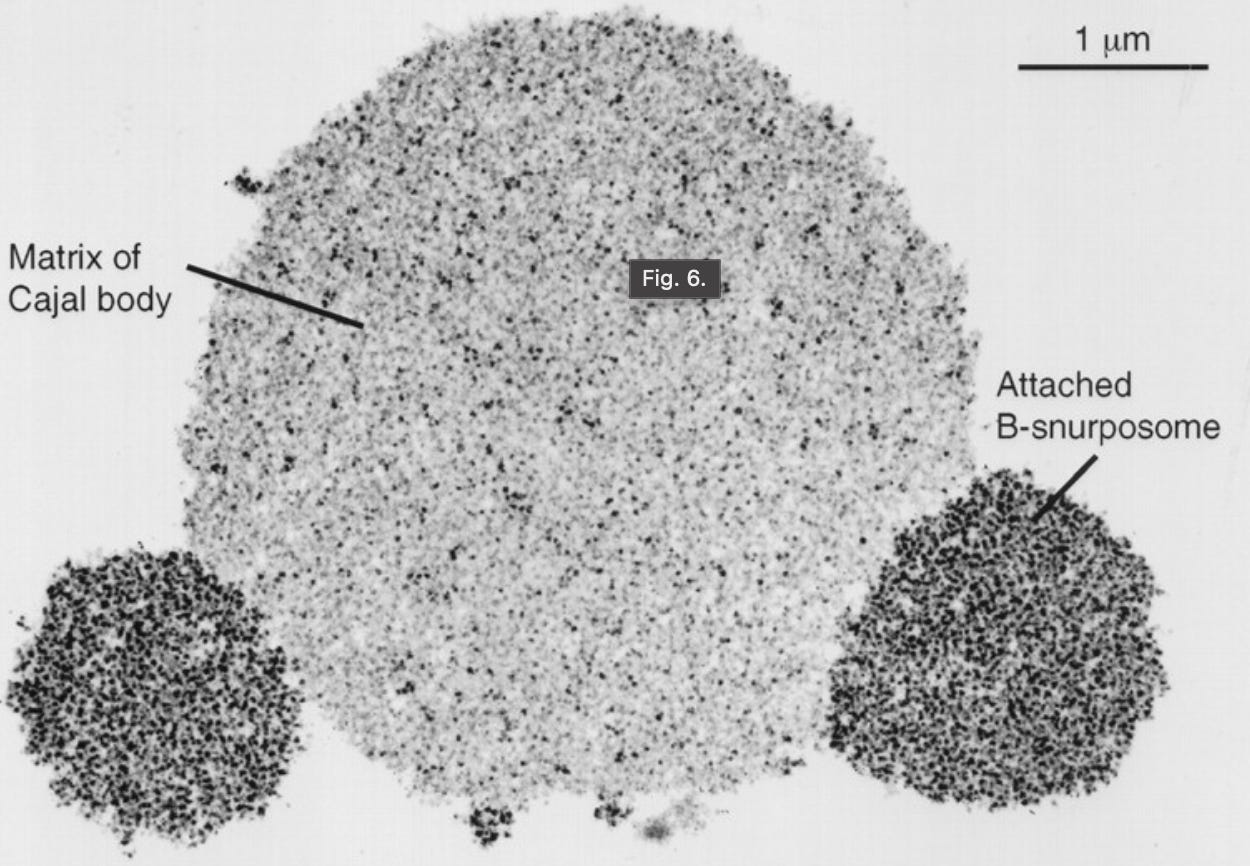
- Cajal bodies, identified by Santiago Ramón y Cajal in 19038.
(BTW, Cajal and Golgi shared a Nobel Prize in Physiology or Medicine in 1906 for research in the nervous system, which is another interesting story)
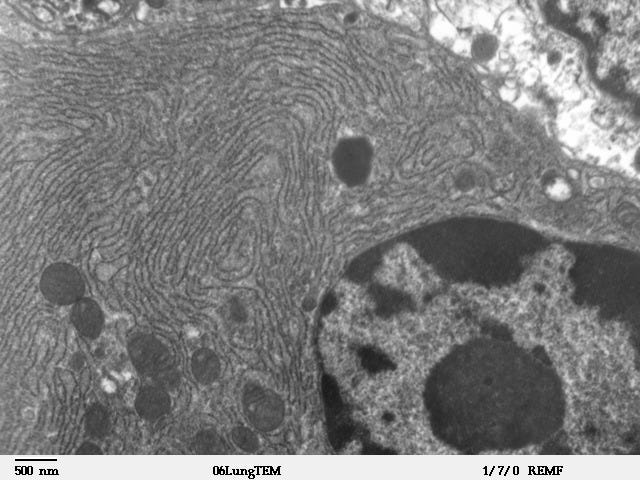
Now we know that some of them (e.g., nucleus, chloroplasts, mitochondria) are membrane-bound, while others (e.g., nucleolus, Cajal bodies) are membraneless.
However, during these early studies, the distinction did not exist, as the resolution of optical microscopes (200-500 nm)9 was too low to reveal membranes (~ 10 nm thickness)10.
It was generally accepted that some form of semi-permeable barrier existed around cells and possibly organelles, but the nature of this barrier remained unclear. It wasn’t until a series of pioneering experiments around 1925 that the lipid membrane nature of cell boundaries was suggested11, 12 (PS: from membraneless organelles, we will see boundary is actually not necessary for selective permeability).
With the use of electron microscopy in 1945, we finally saw the existence of cell membranes13.
At the same time, many organelles were found to be membrane-bound, and detailed structures were revealed. For instance, Porter et al discovered the lacy membrane structure of ER13, thus Porter coined the term "reticulum", meaning "network", which is used until now7. Since then, a membrane-centric understanding of cell models many of us were taught began to take place.
The discovery of membraneless organelles did not stop because of the dominance of membrane-based studies. In the late 20th century, scientists continued to observe structures that lacked membranes. During this time, they often refer to these membraneless entities as “granules”, because the term “organelle” was kinda "reserved" for membrane-bound structures. Examples include:
- Stress granules (1983), discovered in response to stress conditions14.
- P granules (1983), identified as germline-specific granules in C. elegans15.
They also had many other names, such as "bodies", "subdomains", "speckles" et cetera. Only in recent years have they been classified as "membraneless organelles".
Speaking as a biologist, membrane-bound and -less organelles are essentially the same in functions. Both compartment tasks, making "breakrooms" in cells. For instance, you will find abstract structures in the processing of nascent RNA16 is very akin to the processing of polypeptides/proteins17, except that the former is membraneless, and the latter is membrane-bound.
The main difference lies in their physical nature. While the form and function of membrane-bound organelles are relatively straightforward, it wasn’t until recent years that the mechanism behind the formation of membraneless organelles - liquid-liquid phase separation (LLPS) - became clear.
The Science Behind Liquid-Liquid Phase Separation (LLPS)
So, what exactly is liquid-liquid phase separation (LLPS)?
To be clear, the term originates from physics, where "phase" refers to a distinct state of matter, like the difference between liquid water and solid ice.
In liquid-liquid phase separation, two liquids that were initially mixed begin to demix, separating into distinct phases, much like oil and water, or organic and aqueous (water) phases in liquid-liquid extraction. The concept has been studied for centuries, particularly in the context of colloids and emulsions18. These studies led to inventions like dishwasher detergents and laundry powders that emulsify oils and other stains. The term "phase separation" was officially used in the 1940s to describe the behavior of polymers and mixtures, first notably introduced in the paper by Flory in 1953, titled "Principles of Polymer Chemistry"19.

Despite its origins in physics and materials science, biologists have long been familiar with the concept. After all, many studies in colloids and emulsions were with biological materials. For example, milk is a classic emulsion of proteins dispersed in water, showing properties of both colloids and solutions, thus classified as hydrocolloids20. From a physics perspective, you could even consider membranes as a special case of liquid-liquid phase separation, where lipid molecules demix from water solutions into a specific bilayer structure.
During the late 19th and early 20th century, in studies of the physical properties of cells (notably, membranes were still invisible at the time), scientists saw cells as a kind of "jelly" or "colloid", and have noticed the phase-separating properties of intracellular structures. Key observations include:
- Otto Bütschli described protoplasm as having an "alveolar" structure in 1894, suggesting that phase separation might be a fundamental organizational principle in biological systems.
- William Bate Hardy22 and Edmund Beecher Wilson23 described the cytoplasm (then referred to as protoplasm) as a colloidal substance.
- WB Hardy linked the formation of biological colloids with phase separation in his studies on globulins, stating that globulin particles dispersed in a solvent could form an internal phase24.
These works laid the groundwork for understanding "phase" in biological contexts, where the cells were conceptualized as a disordered but dynamic system. Later, studies in this area are more about polymer sciences, and biologists' attention to intracellular spatial organization drifted towards membrane systems, which are more ordered, structured, and easier to understand, until.......
The Renaissance of Phase Separation in Modern Cell Biology
The tipping point was in 2009, when Anthony Hyman and Clifford Brangwynne demonstrated that P granules in C. elegans formed liquid-like droplets via LLPS25. Again, the fluid nature, the phase separation within cellular spaces were reconsidered.
Since membrane theories had dominated cell biology for decades, there was initial skepticism around LLPS since then. After all, under the right conditions, many proteins and RNAs can phase separate in vitro. How relevant is this to REAL biology?
However, the concept gradually became widely accepted as more examples of naturally occurring LLPS were discovered. In my opinion, the strongest evidence came from conserved membraneless organelles found in all eukaryotes, including:
- Nucleolus, involved in ribosome assembly and maturation.
- Cajal bodies, involved in RNA splicing, ribosome biogenesis, and telomere maintenance.
- Nuclear speckles, sites for splicing factor storage and modification.
Now the mechanism of LLPS has been well-studied. The LLPS process is driven by interactions between biopolymers. These are usually multi-valent interaction, such as oligomerization, binding among repeat units, or the condensation of intrinsically disordered regions (IDRs)26. In general, LLPS occurs when specific biomolecules - such as proteins and RNA - exceed a critical concentration and demix from their surrounding environment (cytoplasm or nucleoplasm) to form highly-concentrated liquid-like droplets. Given the process, a new terms "biomolecular condensate" is coined to refer to those membraneless organelles as well.
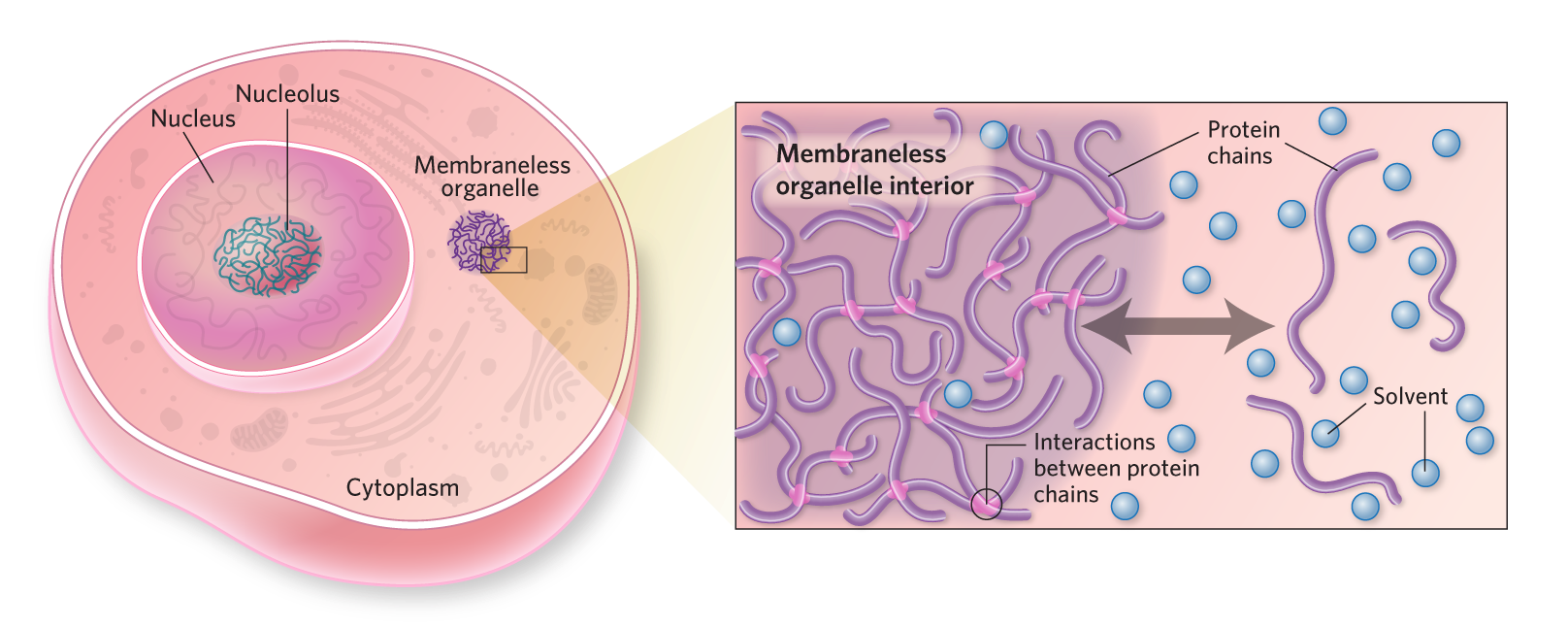
LLPS is sensitive to post-translational modifications, the physiochemical environment, and mutations. These liquid droplets can assemble and disassemble rapidly in response to cellular signals like stress or temperature changes. Mutations can lead to abnormal phase separation, causing diseases such as neurodegeneration.
These droplets are not random. They serve important functional purposes by compartmentalizing reactions without needing a membrane, but still maintaining selective permeability, actively exchanging materials and information with their surrounding environment. Complex structures could also occur, such as multi-layer droplets28, droplet assemblies29, and meshed networks30.
Why is LLPS a Great Design Principle for Synthetic Organelles?
To appreciate why LLPS is ideal for building synthetic organelles, let’s first consider the limitations of membrane-bound structures. Constructing a membrane-bound organelle from scratch is extremely complex. Simply adding new lipid molecules to an existing membrane-bound structure requires numerous enzymes27. Building a vesicle from scratch is even more difficult, as the mechanism remains unclear to this day. Even if we manage to build a vesicle, we still need to add functional pores and receptors to enable selective permeability. The complexity is immense.
In contrast, LLPS offers several advantages for synthetic organelles:
- Simplicity: LLPS can occur with a single molecule, making it even simpler than building protein-shelled bacterial microcompartments, which often require multiple components.
- Engineering: We have plenty of tools to engineer RNAs and proteins, allowing us to easily modify phase-separating molecules and program their functionalities.
- Flexibility: LLPS structures can form and dissolve as needed, in various sizes and shapes, without rigid constraints.
- Energy Efficiency: LLPS relies on molecular interactions, so it doesn’t need the energy-intensive machinery required to build and maintain membrane-bound organelles.
In short, LLPS offers a straightforward, adaptable way to create de novo cellular compartments - an ideal platform for building synthetic organelles.
Building Synthetic Membraneless Organelles: From Concept to Reality
The discovery of LLPS has opened up a whole new frontier in synthetic biology. By mimicking nature’s membraneless organelles, we can build custom structures that are more dynamic, flexible, and efficient than traditional membrane-bound organelles. In fact, to study the properties of phase separation, many engineered phase-separating proteins have been designed to build model systems in vitro26.
The next step? Making it happen in vivo! In Chapter 3, we’ll dive into the design principles behind synthetic membraneless organelles, review successful designs to date, and explore what we can do to improve them in the future. Stay tuned!
Reference
- https://en.wikipedia.org/wiki/Cell_nucleus#History
- Pederson, Thoru. "The nucleolus." Cold Spring Harbor perspectives in biology 3.3 (2011): a000638.
- Staehelin, L. Andrew, and Dominick J. Paolillo. "A brief history of how microscopic studies led to the elucidation of the 3D architecture and macromolecular organization of higher plant thylakoids." Photosynthesis research 145.3 (2020): 237-258.
- Kölliker A. Einige bemerkungen uber die endigungen der hautnerven und den bau der muskein. Zwiss Zool. 1857;8:311–325.
- Dröscher, Ariane. "Camillo Golgi and the discovery of the Golgi apparatus." Histochemistry and cell biology 109.5 (1998): 425-430.
- Garnier, Charles. The basal filaments of glandular cells. Preliminary note . Berger-Levrault, 1897.
- Porter, Keith R. "Observations on a submicroscopic basophilic component of cytoplasm." The Journal of experimental medicine 97.5 (1953): 727-750.
- Cajal SR (1903). "Un sencillo metodo de coloracion selectiva del reticulo protoplasmico y sus efectos en los diversos organos nerviosos de vertebrados e invertebrados". Trab Lab Investig Biol Univ Madr. 2: 129–221
- Abbe, E.K., Beiträge zur Theorie des Mikroskops und der mikroskopischen Wahrnehmung, Archiv für Mikroskopische Anatomie (1873) vol. 9, iss. 1, pp. 413–68, DOI: 10.1007/BF02956173.
- Hine, Robert. "Membrane." The Facts on File Dictionary of Biology. 3rd ed. New York: Checkmark, 1999: 198.
- Lombard, Jonathan. "Once upon a time the cell membranes: 175 years of cell boundary research." Biology direct 9 (2014): 1-35.
- Gorter, Evert, and F. J. E. M. Grendel. "On bimolecular layers of lipoids on the chromocytes of the blood." The Journal of experimental medicine 41.4 (1925): 439.
- Porter, Keith R., Albert Claude, and Ernest F. Fullam. "A study of tissue culture cells by electron microscopy: methods and preliminary observations." The Journal of experimental medicine 81.3 (1945): 233.
- Nover, L., K. D. Scharf, and D. Neumann. "Formation of cytoplasmic heat shock granules in tomato cell cultures and leaves." Molecular and cellular biology 3.9 (1983): 1648-1655.
- Strome, Susan, and William B. Wood. "Generation of asymmetry and segregation of germ-line granules in early C. elegans embryos." Cell 35.1 (1983): 15-25.
- Gadal, Olivier, and Ulf Nehrbass. "Nuclear structure and intranuclear retention of premature RNAs." Journal of structural biology 140.1-3 (2002): 140-146.
- Cooper, Geoffrey, and Kenneth Adams. The cell: a molecular approach. Oxford University Press, 2022. "Protein Folding and Processing"
- https://en.wikipedia.org/wiki/Phase_separation
- Flory, Paul J. Principles of polymer chemistry. Cornell university press, 1953.
- https://en.wikipedia.org/wiki/Colloid
- Butschli, Otto. Investigations on microscopic foams and on protoplasm. 1894.
- Hardy, William B. "On the structure of cell protoplasm: Part I. The Structure produced in a Cell by Fixative and Post-mortem change. The Structure of Colloidal matter and the Mechanism of Setting and of Coagulation." The Journal of physiology 24.2 (1899): 158.
- Wilson, Edmund B. "The structure of protoplasm." Science 10.237 (1899): 33-45.
- Hardy, W. B. "Colloidal solution. The globulins." The Journal of Physiology 33.4-5 (1905): 251.
- Brangwynne, Clifford P., et al. "Germline P granules are liquid droplets that localize by controlled dissolution/condensation." Science 324.5935 (2009): 1729-1732.
- Bracha, Dan, Mackenzie T. Walls, and Clifford P. Brangwynne. "Probing and engineering liquid-phase organelles." Nature biotechnology 37.12 (2019): 1435-1445.
- https://en.wikipedia.org/wiki/Flippase
- Lafontaine, Denis LJ, et al. "The nucleolus as a multiphase liquid condensate." Nature reviews Molecular cell biology 22.3 (2021): 165-182.
- Gall, Joseph G., et al. "Assembly of the nuclear transcription and processing machinery: Cajal bodies (coiled bodies) and transcriptosomes." Molecular biology of the cell 10.12 (1999): 4385-4402.
- Ma, Weirui, et al. "In vivo reconstitution finds multivalent RNA–RNA interactions as drivers of mesh-like condensates." Elife 10 (2021): e64252.

.jpeg)
.jpeg)

.jpeg)